|
����Ŀ�ѧ������ʹ������������̫�ղ���������������������������Դ��һ����ȴһ��Īչ����������ʽ��谭���Ƕ����������ѧ������������֪���������ı��ʺͽ������ȵȣ�������Ϊֹ����ѧ��Ҳ����˸�ʽ��������������Դ�����ۻ���裬���ǿ�������Ī�⡢�ͷ�������ȴ�����ǰ����������Ϊû��һ���ܾ��������ܵ����á���
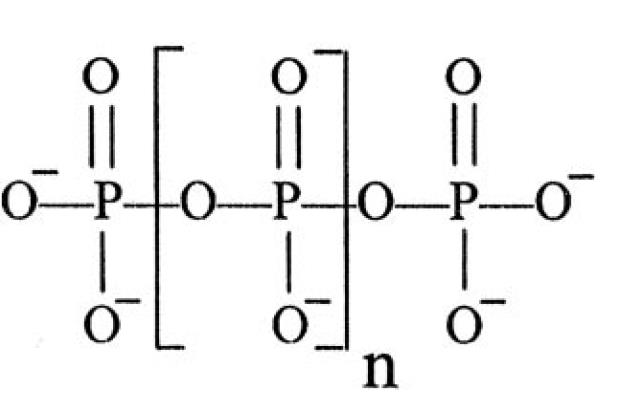
����Դ�Ժη����������������Դ��������һ���⿴�Ƽ���ʱ���������������Ȼ֪֮���٣�����˵����Ȼ�����˲�и��Ŭ��������Զδ��������ŷ��Ĵ𰸡������ﲻ�����ϵ۴����ۣ���Ϊ�ⲻ�ǿ�ѧ����һ���ڽ̣�Ҳ���������������ۣ���Ϊ����һѧ˵����һ�������������棬�������������е�����������β������أ���ˣ��������ڽ��ܹ��ڷ����ڵ�������ϲ���ԭʼ�����Ļ�ѧ����;����صĸ��ּ�˵������RNA�����˵�����߽�����һ�������е����������������Լ��ļ��⡣����Ϊ��ÿһ�ּ�˵�����������ڽ�ʾ������һ�������һ���㣬��û����һ���ܻ���һ����ͼ������ʮ�ִ�������߸��ҵĸо�����ä������ 1.��ԭʼ������˵��"Soup" hypothesis�� ������Դ��ԭʼ�������л���ġ�ԭʼ����primeval soup������˼�������Ӣ������ѧ�Ҵ���ģ�CharlesRobert Darwin�����������ﻯѧ�Ұ°��֣�Alexander Ivanovich Oparin���ȣ����ǣ�ʵ���Ե�֤��ֱ��20�������ڲſ�ʼ���֡�1952�꣬������ѧ�����գ�Stanley Lloyd Miller��ģ��������������ڳ����£���һЩ�����ӣ��ɹ��ϳ��������л����ӣ��簱���ᣩ������ʵ��֧���˰°�����20���ͳ�����Ĺ���������Դ��һЩ���룬�������ڵĵ����ϣ����ܴ���һ����ԭ�Ե�ԭʼ����Ȧ����Ҫ��CH4��NH3��H2O��H2�ȣ��Լ�ԭʼ�ĺ����������ý���Щ����ۺϳɶ��ְ����ᣬ��ʹ�ڳ��³�ѹ�£���Щ������Ҳ�п����ھֲ�Ũ�����ٽ�һ���ݻ��ɵ����ʣ��������ʺ������Ķ�������߷���֬�࣬��һ����ʱ����п��������������� �ѹʵ�������ѧ��RobertShapiro��1935�C2011���� ��Origins: ASkeptic's Guide to the Creation of Life on Earth��һ���ж�ԭʼ����˵�������ܽ�����ڵĴ��������ǻ�ԭ�Եģ��ڻ�ԭ�Դ����ڱ�¶�ڸ�������������²����˼�С���ӻ��������Щ������Ũ���ɡ�����������ܷ����ڲ�ͬ�ĵط���������������Һ��ڵȣ�����ͨ����һ����ת������Ϊ���ӵ��л��ۺ������������������л����оͷ�չ�����ˡ��١�����ͨ��ʵ��õ���֤ʵ�������Ǣ���γ�Ϊ��ʵ��Ȼ��һ�����ţ�Shapiro 1987���� ������Ϊ�����ռ���������Ƶ�ʵ���о��ܹ������ڸ����㹻�����������£��Ӽ��л��������������������Լ��л��������Ȼ��Щ���ɵİ��������������뵰���ʰ����ͬ������ֻ��������Դ���������һ���� 2. ճ����˵��Clay hypothesis�� 20����60������ڿ�ʼ��Ӣ��������ѧ�ҿ���˹•ʷ��˹��Cairns-Smith A. G.��������һ�ֳ�Ϊճ����˵��������Դѧ˵�����ڡ�Genetic Takeover: and the Mineral Origins of Life��һ����ָ����ճ����������Һ�е����Ҹ��ƿ����ڷ�����������ʺ��л�����֮���ṩ��һ�����м䲽�衣ճ���ڹ�������Һ����Ȼ�γɣ�����������һ����ճ�����������ǵ����������ѡ������������У��������ⲿ��������ʽ��һ���ض��ⲿ��ʽ��ճ�����������һ��Ӱ�����ǽ�һ�����ƻ��������ķ�ʽȥӰ�����ⲿ������Cairns-Smith 1987���� ����˹•ʷ��˹��Ϊ����������沶��һ����ʽ�ķ��ӵ�ճ������Ҳ����һ����Ȼѡ����̣������ڴٽ���DZ�ڵĸ����������൱���ӵ�ԭ�л����ӣ�proto-organic molecules���ܱ������α������Ծ��廯�����գ�����Щ���ӵķ��Ӵ����ǵľ���ģ�����γɡ��Ŵ��ӹܡ���genetic takeover���ͱ����һ�������ĸ��ƺ��ģ��ڽ����ϵ���һ�̾�����ν�����������Ӧ��the first exaptation������������ΪDNA�γɵ�һ�ֿ��ܽ���ģʽ�� ����������л���������λ�õ����⣬����˹•ʷ��˹��1995����������Դ������������һ������Ϊ��ijЩ������ģ�������û�õģ�����ԭʼ�����Ϸ����ķ�Ӧ�������ִ�ֲ��Ĺ��������������ӡ�����Щ������ȴ���������ij̶���ģ������ֲ��Ĺ�����á��������ߵ������£�ijЩ�ṹ����������ˮ��֮�����ʹ������̼���̲������ϳ������֮���С�л����ӡ�ijЩ���������Ҳ�������ַ�ʽģ�¹������ЧӦ�����̵�Ҳ���л�����һ�ַdz����ӵķ�Ӧ���̡����ָ��ӵķ�Ӧ����ͬ��Ҳ��һЩ�����ʿ������̶��ϼ���ģ�¡���Ϊɰ��һ�ִμ��ɷ֣������Ķ�������Ҳ���й̵���������̫�������䵽�������ʵij�ʪ�ľ���֮�ϣ���ʱ�ͻ��������ĵ�ת��Ϊ��������һ����̬�ĵ��������֮�±Ƚ�����չ��Ϊ�簱����֮��Ľϴ�ķ��ӡ��� ����˹•ʷ��˹��1995����Ϊ����ЩС�л����ӵġ��ϳɵ�Ȼ������������������������ķ�Χ�ڽ��У����ھֲ�������˵���ڳ�ʪ�Ŀ����ʱ�¶�ڴ������Լ����ں��д��������ߵ�̫������֮��ʱ�����ֺϳ�����ȫ��������õ��ġ��� ճ������������Щ�л����Ӷ����������к������أ�����˹•ʷ��˹��1995��˵������������������ϳɵ��л�����֮һ���������ȶ���Һ�����ԣ��Ӷ���������ճ���ᾧ���̡��ȼ�����������һЩ�IJ���Ҳ�൱���ϳɡ����֪���������ܰ���ճ���ϳɡ����м��ַ���ͨ�����������ܽ�Э��ճ���ϳɡ��������Լ�С���İ����ᣬ��������һЩ���ӣ������ڿ�����Һ�еĽ������ӡ������������ĸ����һ������ճ����ճ����֮��ķ��ӡ����������г��ֵ����������ε�λ���ر��ʺ���ճ����ı�Եճ�ᡣҲ�����������ַ������Ⱦ���Ϊ����ճ������ö���Ƶġ�����������������һ����ǵ�λ�ϳɡ�����ճ���л����������������Һ�����ܵ��������������ɻ��߱�ˮ�����������������ʱ�����ǵ���;�ͻ���ȷ����ʾ������Ҳ���������Ǻ���ͺ��Ǻ����ǰ������һ��������ǡ������Dz��ǣ��㶼�����������⣺���������Ǻ������Ǻ����ǰ���ڿ�ʼʱ���������Ŵ���;������������ӿ��Ը�����Ϣ�Ǻ��������飬�������ַ�����һ��żȻ�¼�����ij�ֶ���Ϊ��һ��Ŀ�Ķ������������ȴ������ͨ���������ֽ�����Ķ�������������;���������̸����ʲôԤı����ֻ��һ�ֻ������塱�� ���߲�����ճ����Ϊƽ̨��ģ�Ӷ�ԭʼϸ�������Ŀ������壬Ҳ�����Ͼ����������ԭʼ�����������С�����л���������ۻ��еĿ������á����ǣ�����Ϊ��Ҫ������ʼ�أ�����������ճ�������е����������ʡ�RNA��DNA�����������Լ����չ��ɵ�ϸ���أ���Щ����ճ��ѧ˵�����ش�����⡣ 3. ֬�������˵��Lipidworld hypothesis�� ��ɫ��ѧ��Daniel Segr�����������ν֬�������˵��The lipid world������Ϊ������Ը�����������֬�lipid-like��(Segr�� et al. 2001)���ڽ��������£���֬��ˮ���γ�֬��˫��ṹ����ϸ����Ҳ��ͬ���Ľṹ����Щ���������ڵĵ����ϲ������ڣ����������Եij�������Ҳ���γ�Ĥ�����⣬��Щ֬����ͨ��֬�ʷ��ӵIJ�����ܻ����ţ��ڹ�����չ������¿��ܻ��Է���һ��Ϊ�������屣��ͬ���������֬����ɡ���������Ϊ��֬����ķ����������Ϣ����ij�����ʽ������������RNA��DNA��������Ч������Ϣ�ľۺ���ij��֡� �ڸü�˵�У�֬��˫��Ĥ�ɸ��ָ��������Է��ӻ������϶��ɣ������Ĥ����Щ�������������Ϸ�ʽ�Ǿ�ģ�����ЩDZ�ڵ�����У�Ĥ���ض�������Ӧ���������ڰ�������ν��ѭ��ģʽ������Ĥ��λ���벶������Ĥ�е��ض�������֮����ڻ��������������ϵ������λ��/�������Ӧ��ϵ���Դ����Ӵ�����Ĥ�������ܹ����ܴ������Ȼѡ�����Ĥ�IJ�ͬ��ϵ�ij��֡� ����Ҳ��Ϊ����յ�֬��Ĥ�ij�����ͨ��ϸ������Ҫһ�������ֽṹ�γɵ��Է���ҲΪϸ������Ȼ�����춨����Ҫ�Ļ�����ͬʱ��Ĥ������������һ��Ϊ��ҲΪϸ�����ѻ��Ƶĵ����춨��һ�����������ǣ�����֬����ķ���������ȥ�Ƶ���Ϣ���������չ��RNA��DNA�ij����أ������۽��ð����ij�ѭ��Ҳ�������µġ���ˣ���Ȼ֬�ʶ�ϸ��������ij���ʮ����Ҫ��������������Ϊ���ĵ�ԭ�� 4. �������μ�˵��Polyphosphateshypothesis�� �������ﻯѧ��MichaelR. W. Brown��Arthur Kornberg���Լ���������ѧ�͵���ѧ��Matthew A. Pasek�ڷ�����PNAS����������Ϊ������������Ȼ�����Ĵ�����龰���ٵ�һ��������ǰ�����Զ��ĵ�����ѧƽ��ڸ��𰱻���ķ���֮�У�ȱʧ����������ۻ���ij���������������ε��������ÿ��Խ����һ���⣨Brown & Kornberg 2004��Pasek 2008����������������ͨ�ĵ�����������PO4−3�ۺ϶��ɣ��м��ֿ��ܵĻ��ƣ������������𰱻���ת��Ϊ���ĵĶ�ۻ�������Ҳ�Ǻϳ���ATP�����Ĺؼ��������ʵĺϺ�����ǰ�壬һ���ؼ��Ĺ۵��ƺ��ǣ���������������η�Ӧ�γɲ���������ƣ���ʯ������˱�����ij�ֿɿ��Ļ�����ά�ָ����Ӳ����������εij����� ������Ϊ���������μ�˵��ע����������������Է��γɵ��������������ɷ��ϣ����ܵĶ�����������Ȼ�γ���ATP�����������������ʵ���Ҫ���������������Ļ�ѧ����������Ҫ���壬����Ҳֻ������ѧ��������֮һ�� 5. ��ø�����˵��Coenzymeworld hypothesis�� ��������Ŵ�ѧ��Alexei A. Sharov����������ѧRichard Gordonͨ����Ħ��������������������Լ����Ʒ���Ϊ��������Դ��97��25����ǰ���ȵ����γ�������ʮ���꣨Sharov 2006��Sharov & Gordon 2013��������������У�����֤�ݱ���������ÿ��3.76���귭һ����������ľ������������ֱ�ʾһ��������������Ҳ���������︴���ԣ�Ʃ��������й����Է�������DNA�ij��ȣ������㡣���������ת���ĸ�������������ϵ����Դʱ����ͼ���õ�һֱ�߹�ϵ����ֱ����X���ཻ�㣨��1�������ᣩ��ʱ��Ϊ97��25����ǰ����Ϊ̫��ϵ������Ҳֻ��46���꣬�����˵�����������ǴӸ��ϵ�������ij�ַ�ʽ��������ġ� һЩѧ���������۵����һ��˵���������������������ڵ���ȱ������������������ʿ���Ҫ��ö࣬��������Դ�ڼ����������Dz�������ġ�Ϊ�˻�Ӧ��Щ������Sharov�����һ�������Ƹ�ø���ӣ��������뵥�������൱���ļ������Ȼ�����龰��Sharov2009��Raffaelli 2011����������Ϊ������ͳ�ƹ����Թ�Ȼ��һ���Ŀ�ѧ��ֵ������˵��������Դ�Ļ��ƻ������Զ�� 6. �����������˵��Iron�Csulfur world hypothesis�� �¹���ѧ��Wächtershäuser G.��1990����PNAS�Ϸ����ˡ�Evolutionof the first metabolic cycles��һ�ģ������һ����֮Ϊ�����������������Դ��˵����Ҳ��һ����Դ��Դѧ˵��Ҳ��������Դ�ں�����Һ��ڵļ�˵һ�¡�Wächtershäuser��1990����Ϊ�����ƶ�������Դ����Դ��������������������ʣ��������������Щ���������������ԭ��Ӧ���ͷŵ����������������л����ӵĺϳɣ����ҿ����ڵ;����������γɣ���������ϵͳ���ܽ����������ܹ����Ҹ��Ƶ��Դ�ϵͳ�Լ�������������֪��������ʽ֮ǰ���ܹ��³´�л�Ļ�Ķ���ʵ�塣WächtershäuserҲ�Ǵ�л�������ߣ���Ϊ��������Դ�д�л�����Ŵ�������Ĵ�лָһ�ֲ��ܻ�ѧ��Ӧ��ѭ�����������������ܱ��������������ã�����Ϊ��һ��һ��ԭʼ�Ĵ�лѭ���������������Ϳ�ʼ���������ӵĻ���� ������Ϊ�����ֺ�����Һ��ڼ�˵��Ҫ���ٵ�һ�������ǣ���ʹ��Щ���ﵮ���ˣ����ǣ����������Ż�����ý���ãã��ʱ����������ܹ������ȥ����Ϊ��ʱ�������������������Դ��������������Ѿ���ʧ����������ܹ���Ӧ��ȫ��ͬ�����滷����Ҳ�����˻���Ϊ���������ĵ�����������Щ��ڴ���ԭʼϸ�����ܻ���һЩ�����ܹ��ɹ��ݱ�Ϊ��Ӧ��һ�����滷��������Ҳֻ��ͣ����һ����֤ʵ�ļ�������֮�С��ں��滹Ҫ̸��������H2�����ڼ�˵Ҳ����ͬ�������⡣ 7. п�����˵��Zn-World hypothesis�� �¹�������ѧ��ArmenY Mulkidjanian�����һ����Ϊп�����������Դ��˵��Mulkidjanian 2009������ѧ˵��Ϊ��Һ�зḻ��H2S��ԭʼ�������ˮ�ء�����ˮ����õ����˽�����������ij����������Ϊ�κ������ں�������Һϵͳ�ֲ��д�״�ṹ����Ҳ��ӳ�˹��ϵĻ�ɽ����Ŀ�״�������VMS������Һ��Դ�����ֽṹֱ���ɴ������������̫���棬��ḻ������ΪFeS2��CuFeS2��ZnS������һЩPbS��MnS��ZnS��MnS���д�����䣨�������ߣ������Ķ����������ڸ��Ʒ�����Դ��ص�ʱ�䴰�ڣ�ԭʼ����ѹ��Ҳ�㹻�ߣ�> 100 bar���Ա��ڵ���������ZnS��������������Ҫǿ10-100������ZnSý��Ķ��صĹ���������Ծ������ṩ��������Ϣ�ʹ�л���Ӻϳɺ���ĺ˼��ѡ����ʵ������������� ���ǣ�Mulkidjanian�ȣ�2012��ע�������ϸ����������ɶ����ִ���ˮ����ֲ�ͬ�����ǽ�ϵ���ѧ�����Լ��ִ�ϸ���ձ�������ӳɷ��������ϵ�������ع��������ϸ���ġ������������Ʋ��ձ��ԭʼ�ĵ����ʺ���ϵͳ��ʾ����K+, Zn2+, Mn2+�������ε����Ժ���������ѧ�ؽ���ʾ��������ϸ����Դ����������ɲ��������ڽ������dz�ν�ĺ��ף��������ǽ����ν����½����ϵͳ����Ҫ��ɢ�������ĵش�����һ�¡���ȱ���ҳ���CO2��ԭʼ���������£������ȳ���ˮ��������������ѧ�ɷֽ��������ִ�ϸ�����ڲ���������ˣ�������ǰϸ���ڿ��ܷ�����dz�ġ������ˮ�ء������������Ŷ�Ĺ����ο����ʣ�����Ž������������K+, Zn2+����� ��ѧ˵�����ִ�����ϸ���еĹؼ�����Ũ�ȵı�������ԭʼϸ������֮�ؽ������Ʋ⣨������ֻ�Ƕԡ������ˮ�ء���˵�����䣩��������Ϊ�����ɵ�ϸ����û�����������ӵ�ϸ��Ĥ��Ҳû��Ĥ���ױã���ôֻ���뻷������ƽ���ˣ�����ִ�ϸ���е����ӹ���Ҳ����ӳ��ԭʼϸ����Դ�ص������ӹ��ɡ����ǣ���β��ܱ�֤��һ�����dz������أ� 8. ����������˵��PAH world hypothesis�� һЩѧ������������Դ��һЩ�������Ȧ�⣨�Ǽʣ��ĸ��ӷ��ӣ���Ϊͨ�����������֣��л����Ӵ��������Ǻ�����֮�ϡ���������ѧ��Adolf N. Witt ����ϵ�����������polycyclic aromatichydrocarbons��PAHs����Witt 2003�������������廯ѧ��Garc��a-Hern��ndez�ȣ�2010�����Ǽʼ���PAHs����ϩ��PAHs������Ϊֹ���Ǽʿռ䷢�ֵ���ӵķ��ӡ� �������˹Ƥ�߿ռ���Զ���ӻ���һ�������γɣ���̫ͬ�������γɵ�������������ĺ���HH 46-IR��Բ����������м��dz���Χ�ķ��ӣ������軯�����CO��2012�꣬NASA�Ŀ�ѧ�ұ�����PAHs���Ǽ����ʵ������·�����ת����ͨ���⻯���á��������ú��ǻ������ã�����˸�Ϊ���ӵ��л���ֱ�����ͨ�������ʺ�DNA��ԭ���ϡ�������ͺ�����������һ����Gudipati & Yang 2012���� ������Ϊ��һ��˵һ�����ṩ���Ǽ��д��ڵ��л�������������͵Ŀ��ܣ���һ����Ҳָ������ռ�����л������γɵ���������������Կ�����������ʵ���̫����֤�ɣ������˶��ѣ�����˵��������Դ�ľ�����̡� 9. ��л����ģ�ͣ�"Metabolismfirst" models�� ������ģ�ͷ������"naked-gene"�����Ҹ��ƵĹ۵㣬���ٶ�ԭʼ��л�ȳ��֣����Ϊ����RNA���Ƶij����ṩ�������ں�������Ϊ������������������ѭ���������Լ��ڸ����л������������ϳ��еĶ�����̼�������ӣ�����������ͺ����ᣩ���ƶ�����ʾ���Ǵ�л�������������֮һ��Lane 2010�� ��Smith��Morowitz��2004����Ϊ�������ֵ�Ũ�ȳ��㣬ѭ������������ת������ÿ���м����Ũ�ȵ����������ͻ��Է����л�����һ���м�����ˣ��������㲻�ǻ���Ĵ��죬��ֻ������ѧ�ͻ�ѧ���ﱾ��������ѧ��Carroll��Ϊ����������Ŀ����Ϊ�˻�ԭCO2������л���ȶ������Ŵ������龰��һ���֣���Musser 2011���� ������Ϊ����һ������Ȼ��ʮ�ֲִڣ�����������м�ֵ�Ĺ۵㣬��������������Դ����ν�ġ���������ڡ�������Ŀ����Ϊ�˻�ԭCO2��������˵������Ȼû�н�ʾ�����ı��ʣ������ٱ���Ŀ����Ҫǿ�� 10. �Դ���˵��Autocatalysis hypothesis�� ������������ѧ�ҡ��������۵��������Stuart A.Kauffman�ڡ�The Origins ofOrder: Self-Organization and Selection in Evolution��һ����������������Դ���Դ���ѧ��Ӧ���磨Kauffman 1993������ν�Դ�ָ�ܴ������γɵ����ʣ����Ҳ���мķ��Ӹ����ߵ����ԡ�Rebek�ȹ����˰������գ�amino adenosine�������������pentafluorophenyl ester�����Դ�������������������AATE���Ļ��ϵͳ������һ������ʵ���ϵͳ�����˶��������д������ϳɵ�AATE����ʵ��֤�����Դ�������һ��������Ⱥ�IJ�ͬ�Ŵ�ʵ�����о����Ŀ����ԣ���ɽ���Ϊһ�ֳ�������Ȼѡ��ʽ��Dawkins 2004���� ������֮�������밬�����Է�������֯Ϊ�����ij�ѭ�����۴�ͬС�죬û�б��ʵIJ�𣻴��⣬û�и��루ϸ��Ĥ����ϵͳ����ν��������Ȼѡ��Ҳ�Dz����ܷ����ġ� 11. ����֯���Ը��Ƽ�˵��Self-organizationand replication hypothesis�� ����֯���Ը��Ƴ�����Ϊ������ϵͳ�ı�־������Ҳ�з���������ں��ʵ������³��ֳ��������Ե��������ӡ�Ʃ�磬����ֲ��ѧ��William Martin��Michael J. Russell��Ϊ���ӻ����е�ϸ��Ĥ�������ָ�������֧�ŵ�������ԭ��Ӧ������֯��������ص����ԣ���ˣ������������Ե���������п������������Ĺ�ͬ���ȣ�Martin & Russell 2003����������Palasek��2013��ָ����������Һ��ڵ��������ӣ�RNA�������Է��ؽ���������װ�����������ڲ���������װ���������Դ���о�Ҳ�н�����壬��Ϊ��ʹ��������ʼ��������װ���л����ӵ����۸�Ϊ���ţ�Nussinov et al. 1997��Vlassov et al. 2005��Koonin et al. 2006���� ������Ϊ����ѧ˵ָ��ϸ��Ĥ�����������Լ�����֧�ŵ�������ԭ��Ӧ���������Ĺ�ͬ���ȵ�������һ����ʵ�Ƕ��ж�ԭʼϸ������һ��˵���������Բ���Ϊ����RNA������װ����������Դ���ڵ�����������ȷ�ϣ���Ϊ�����ִ�������˵��û������������Ҳ���������ڡ� ������ѧ���ڡ��������������ղ������Phillip E.Johnson��1940-����Darwin On Trial�����ģ����д���ģ�һ����������������Ȼ��ʵ����ȡ�óɹ���ǰ;���ѣ���Щ��ѧ�Ҹɴ�ʹӼ����ģ�ⷽʽ���֣���������ʵ���Ե��ϰ�����һ���ѧ���Ѿ��ٶ������������Է��������ԺϽ����������ӵĽṹ��������ˣ���ô���ﵱȻ��Ҫ���Է��Ե�������������ˡ����������ּ�˵����ѧ�ҿ������һЩ����ģʽ��ģ��������Դ����������������ģʽ����ʵ�кι���������ȫ����һ�������ˡ���Johnson 1991���� 12. ���Һ����Ȧģ�ͣ�"deep-hot biosphere"model��
20����70����������µ�������������ѧ��Thomas Gold��1920 �C 2004)����������Һ����Ȧģ�ͣ���Ϊ�������������Դ�Եر��������ڵر���������֮�£�Gold 1992��2001����20����90������������ʯ�з������������ϸ����С��˿״�ṹ�������ܺ���DNA�����ܿ����Ƕ�Gold���۵�֧�֡� �����Ѿ���֪���ӵر���֮��5���ﶼ���ڷḻ�������Ҫ���ȼ��Ĺ�ϸ����������ϸ�������߸���طֲ���һЩ���Ӵ��Ļ���֮�С�Gold���ƣ�����ɼ�֮��ʳ�����ϸ����������˵����Ҫ�ģ���Ϊ��һ��ˮ���е����������ܻὫ���е��л���������һ�գ�����Լ�Ҳ�������Gold��������Ϊ��������ʳ����Դ�ڵ����ԭʼ�����������������Զ�����̼�����ʳ�﹩����ͨ���Ľ������л��忿ˮ����ʯ�еģ���ԭ�ԣ���������֮��������ͷŵ�������ά����� ������Ϊ�����Һ������������ʵ�������������Ƿ�����������֮����������һ���£��Ȳ����ų����ֿ��ܣ�Ҳ��ȷ������ˡ����ǣ���Щ����Ĺ�ͬ����������ܴ����������ĵ���˳��������沢�ɹ�ת���Լ��Ĵ�лϵͳ����Ӧ�µ�ȱ�������H2�ĵر��أ�����ر����������������ʣ��������ֺα������ڵ��¶��������أ� 13. ������˵��Deep sea vent hypothesis�� ���ڻ������Һ��ڼ�����Ϊ������������Դ�ں�����Һ��ڣ������︻��������Һ��Ӻ���ӿ������ʽ8-1����������һ��ϵͳ�У������Ļ�ѧ����������ԭ�������ṩ����������ӹ��壨��H2����������壨��CO2��������Ӧ�� 3Fe2SiO4+ 2H2O �� 2Fe3O4 + 3SiO2 + 2H2 ��8-1�� Martin��Russell��2003����Ϊ��������ڴ�����һ���Է������Ӷ����ƻ�ѧ���ݶȣ�����������������Ȼ���������볡���������ϸ���ң�Ϊ�л����ӵ�Ũ���ṩ��һ����Ȼ�ķ�ʽ����Wächtershäuser�������������Щ�������������ķ�������ɵĸ����������д����ԣ�Lane 2010�������ӵĿ�Ĥ�˶�Դ�������������ӵ���ϣ� �� ��Ũ���ݶ��������ɢ���������������ڵ����п����������ڴӸ�Ũ�����Ũ����ɢ�� �� �ɵ����ݶ�����ľ�������������H+�������������������ص���������ɢ���������������෴�� �������ݶȺ���һ����ǵ绯ѧ�ݶȣ�Ϊ�Է��ĺϳ��ṩ���������Ӷ�����Ҳ�ܱ���Ϊ����Ϊ��Ĥ������Ũ�Ⱥ͵��Ʋ�ͬ�����������ѹ�ݶ�����ϵ����ܵ�һ�������� ������Ϊ������ֻ���������ִ����������������ԣ����⣬��һ����������ǰ��˵������Щ����Ĺ�ͬ�����ڱ�ϡ�͵�ãã����֮������ܹ��ɹ���Ӧ��ȫ��ͬ���µĻ��������⡣ 14. �Ⱥϳ������˵��Thermosynthesisworld hypothesis�� �ֽ͵���������ѧ����ͨ��������ѭ�h��������øA;�������У����߶���ԭʼ���������˵����ϵ�������������ﻯѧ������ѧ��Anthonie Muller������Ⱥϳɼ�˵���ǻ�ѧ������������ѧ���̣�����ϸ��������������а�������Ҫ��ɫ���ȷ���Ϊ������Muller 1985��1995�������Ⱥϳ����磬ά�ֻ�ѧ����ATP�ϳ�ø����Ϊ�����ִ��ø����ӽ�����Ĵ�л���̵ġ� ���ȣ�������Ҫ��Դȥ�ƶ����۷�Ӧ�Բ����������ʵĶ��ļ���RNA�������������һ����˵�����ڽ����ATP�ϳ�ø�Ľ�ϱ仯���Ƶ������仯������һ�������ʡ�Ӧ�ý���˵�����ġ����ᡢ�����ᡢRNA�����塯����������Ũ����һ����Ӧ����������ᱣ�ֽ��ֱ������һ���¶ȱ仯���ű����յ�ȥ�۵����ͷš��Ⱥϳ��������Դ������ѭ��������һ��������ˮ����ԭʼϸ�������Ľ��������һ����ɽ��Ȫ������һ��������������һ������Դģ������Ҫ������֯�ͺ�ɢ�ṹ����ѭ�����ȷ���ϸ�����ѹ�������Ȼ�������ձ��ɫ����Ϊ��ԭʼ�Ⱥϳɵ���� ͨ�����ữϸ��Ĥ֬�ʣ����������ĵ��ס�ʹ���е���֬��ԭʼϸ��������ѡ�����ơ�һ��ʼ���������ĵ���Ҳ�ϳ���һϵ�е��ף�����ֻ�м����������Ⱥϳ���������������Ӣ����ѧ����ѧ��Freeman Dyson�ڡ�Origins of Life��һ�飨Dyson 1985����������ģ��������ĵ������Եط�ֳ�ˣ������������������ĺ��������û�п������������ܺ���ɲ�ͬ�İ�����������ɡ�����������ϳɵ�RNA�У�RNA���и���������ϳɵ��ٶȺ�Ч��������ѡ��Ʃ�������Ϊ��ʹRNA��ת��RNA�������RNA�����ɣ����߸�һ��أ�RNA��������г�Ա��������ѡ�����������������Ⱥϳ�����ɽ����Ŵ���ϵ����Դ�� ����������ȷ����һ�����ѭ��·������˼ٶ���ø�Ĵ��ڣ����ǣ��Ⱥϳ���������û�ἰһ��ͨ·��Ҳû�мٶ�ͨ��ø�Ĵ��ڣ�ATP�ϳ�ø�Ľ�ϱ仯����������һ�����������ܵ������������̣�Muller &Schulze-Makuch 2006����������һ�������ļ��������ܵ�ø�Ļ��ơ�RNA����Ҳ��ζ������ø�Ĵ��ڡ��������ƣ������������һ��ø�����Ǿ������ܵ��£�Orgel 1987���� ����һ�����������ȵļ��裬��������Ϊ�������ͨ����������DAN��RNA��˵���ϲ����������ŷ��� 15. �����Ժ�̲��˵��Radioactivebeach hypothesis�� �о���������ѧ������ѧ��Zachary Adam�����һ�ֳ�Ϊ�����Ժ�̲��������Դ��˵��Adam 2006��������Ϊ��������ijһʱ�������ij�ϫ���̣��������dz��ӽ�ʱ��������ԭʼ�����ĸ�ˮλ��־��Ũ�����˺�����������Ԫ�ؿ��������ܵ����������������ʵ���Դ�������ķ����Ժ�������Ϊ�л����ӣ������ᡢ�ǣ��IJ����ṩ���㹻�������������Ե��������monazite�������ͷ�������õĿ������ס����������ᡢ�Ǻ��ܽ��������ͬʱ�γ��ˡ������ַ�Ӧ��������������һ��Ũ�ȵķ������������γ����л�����������IJ��֣���Щ������������������̵����ڴ����� ������Ϊ��һ��˵Ϊԭʼ��������֮���ṩ��һ�ֿ��ܣ����������������Ϊ����������ϳɼ��л��������ṩ�������������������ڵĴ��������ǣ����������Լ�����ϸ�����������Դ���أ� 16. �����ߺ��¶ȸ����ĸ���ģ����Ultraviolet and temperature-assisted replication model�� �����ߺ��¶ȸ����ĸ���ģ����Ҫ�ǻ���������Դ���ȶ���ѧ�ӽǡ�ī�����о�������ѧ��Nuclear Physics���ͽ�������ѧ��Physics of Evolution����ѧ��Karo Michaelian ��Ϊ���κι���������Դ��ģ�Ͷ����뿼����������ʵ����������һ����������ȶ���ѧ���̣����IJ���������������������أ��صIJ�����������˵������żȻ�ģ������������ڵĸ���ԭ�����ڵ�����ͨ���ٽ�������ɢ��ˮѭ�������ӵ�����ص���������ˣ�����ֽ�����������ѧ�����ǰ���ˮѭ���������أ�������������֮��ҲӦ����ˣ���ʵ�ϣ�ˮ��Һ�е�RNA��DNA��200-300 nm������Χ�������ķdz�ǿ�ҵ������ߣ������ܴ���Ũ���ǰ��������㵽��ر���̫�����һ���֣�Michaelian 2009��2011a���� Cnossen�ȣ�2007����Ϊ����̫����ʱ�ڵ���������IJ�����260 nm����Ҳ��RNA��DNA������ǿ�ҵIJ����������������������ڵ�31�����������������̫���溣�汻�л����ӵ����պͺ�ɢ���������˱����¶ȣ������������ӣ����Ҳ������ԭʼ��ˮѭ������Ϊ���������ӵ����պͺ�ɢ��һ���ص��������̣�Michaelian��Ϊ�ȶ���ѧ����Ӧ�ôٽ��������������е�RNA��DNA�ķ�ƽ�����Ȼ�ϳɹ��̣�Powner et al. 2009���� ��һ�����ؽ���ø�Ϳ��Խ���RNA��DNA���Ƶļ��ƣ���������ͬ��������ѧ������ṩ��������������ʱԭʼ������¶��Ѿ���ȴ���˲��RNA��DNA�ı����¶�֮�£������ϷǰͲ��ٴ�Լ35��33����ǰ�����Ҵ���ʯ�е�18O/16O�ı��ʣ������¶ȹ���ԼΪ70��15 ��C���ӽ�RNA��DNA�ı����¶ȣ���Lowe & Tice 2004���������ϣ�����ˮ�½����͵������¶�����ʱ������RNA/DNA����Ϊģ���γ�˫����RNA/DNA�����˰��죬RNA��DNA�����������ߣ�����ת��Ϊ�ȶ�ʹ�����¶����ߵ���ʹRNA��DNA���ԣ����ֿ�����������ҹѭ���з������У�Michaelian 2010��2011b���������¶ȸ����Ŀ���������ʵ�����ճ���������DNAƬ�ε�PCR���ۺ�ø��ʽ��Ӧ�����ơ�Michaelian��Ϊ����ͳ�Ĺ���������Դ���о��ڴ�ȥ��������ƽ�����������������������Ǵ���ģ����뿼�Ƿ�ƽ����������ر�����������������������Ҫ�ԡ� ������Ϊ��ѧ˵�ṩ��һ��������ӽǣ����ǣ���������ЩRNA��DNA�����������Ϊ�β������أ� 17. ˮ�ݼ��裨Bubbleshypothesis�� ��������IJ��˲������µ���ĭ��ϯ�����������������������������Ϻ������������ֺ����߸�ĭ�뱻�紵�����л��������ں�̲����ã�������dz̲�ϵĺ�ˮ�������ů������ͨ�����������Ũ���л����ӣ���Ҫ��ˮ���ɵ�ˮ�ݺܿ�ͻ����ѣ������������ӵ�ˮ�ݾͻ��ȶ��ö࣬�������еĹؼ���Ӧ�и�����ʱ����С������Ӿ�����ˮ��ͷ������ˮ��β����һЩ��������ˮ�����Է��γ�Ĥ������һ�����κ�ˮ�ķ�յ�Ĥ�����ִ�ϸ��Ĥ�ļ����ǰ�塣���һ����������������ˮ�ݵ������ԣ����ˮ�ݾͻ������ƣ��ᱻ���ڸ��ӽ�����Ȼѡ��ȴ���¼�еĶ��ˡ���ˮ������ʱ������Ϊԭʼ�ķ�ֳ��ͬʱ����Χ�Ļ���֮���ͷš�ʵ�顱�Ľ����һ���㹻 ����ȷ�Ĺ���ԭ�ϡ����ͷŽ��������Ϳ��ܳ��������ԭ�������Լ��������Ͷ�ϸ������ķ�չ��Panno 2005���� Ӣ��������ѧ�ҿ���˹��˹��˹��1995�������������һ�����룺����ȱ��״̬�£�������ӽ����������ӡ������������������һ��ʢ��Ӫ�����ľ���룬Ϊ���������ӡ��Ļ����ṩ�˳������������ɻ�������ɡ�����ֻ�����У����ַ��Ӳ��Ͻ�ϡ����ѡ���Щ����֮��Ľ�ϻ���������ӵĽ�ϸ�Ϊ�ȶ������ǣ����ֽ�Ͼ��γ����������۵�С��ĭ��С���顣����ʱ������Щ��������ķ�ĭ��ɢ��С�飬��ЩС���ĭҲͬ���������µ����ʡ�������ַ��ӽ��������һ����Ҳֻ��Ҫһ�����ɡ����������˴���������Ľ����Ľ��ݡ������Է��ܣ����Խ������������������ ������Ϊ��ˮ�ݼ����ṩ��ԭʼϸ����һ�ֿ���ģʽ����֬�������˵��ԭʼ������˵��������Щ���ƣ���ͬ��δ�����漰�������ӵ��γ����⡣ 18. ����Դ��˵��Multiplegenesis�� �ڵ�����ʷ�ij��ڣ���ͬ��ʽ����������ͬ����quasi-simultaneously�����֣�Davies 2007������������ʽ���ܾ����ˣ������䲻ͬ�����ﻯѧ���̣�Ʃ��������������������ر�Ļ�ʯ������Ϊ�ȼ�������Ҵ����������ؽ��������������ڵ��������ϵ��������ƶ�δ��ע�⡣Hartman��1998��ͨ���������۵�������������¹۵㣺��������������Ҹ��Ƶĸ�������ճ�������̶�CO2������ת���ɲ�����������Ȼ��ᣬ���ţ�������ܸ��Ƶ�ճ�������ǵĴ�л�����ɵ�ϵͳ��������Ҫ�̵������ĸ��������Ȫ������������ϵ�һ���ܺϳɺ��������֬�Ľ���ϵͳ֮�С��������ϳ�����������Դ˵����ô������ĺϳ����������ʺ���़���ĺϳɡ����⣬�����������ۺϳɶ�������ͨ����ۺ�����İ���������ֱ�Ӿۺϡ� �ѹʵ���������ѧ��LynnMargulis��1938�C2011�����ڹ���������Ϊ����ͬ��ʽ�Ĺ�ϸ��ͨ�������γ������ϸ����Margulis & Sagan 2001�����ڲ�ͬ��ϸ��֮���Ŵ����ʵ�ˮƽת�ƴٽ��������Ĺ�����ϵ����ˣ����ͬ���л�����ܲ��빱�����ձ��֮Ϊ�����ִ�����Ĺ�ͬ���ȣ�LUCA���Ĺ����� 19. ��������Դ������� ����ѧ�߰°�������1924�����ġ���������Դ����˵�������ź���ϸ������Դ��Ȼ�Ǹ����⣬Ҳ��ȫ��������������ڰ���һ�㡱��1988�꣬�¹�ѧ�ߵ�˹��Dose K��������һƪ������˵�����ڻ�ѧ�ͷ��ӽ������������������Դ��ʵ��������30���ꣻ��Щ����ʵ����������������⣬��û���ҵ��������İ취��Ŀǰ�����й�����Щ���۵����ۺ�ʵ�飬�������뽩�־��Ƕ���֪�й��ˡ���1998�꣬����ѧ�߰ʹBada J��������һƪ������˵�������죬�����Ǹ��20����ʱ��������Ȼ��Խ���20����ʱ������δ�ܽ�������⣺���������ڵ����Ͽ�ʼ�������ź���2003���� �������ﻯѧ�ұ�ϣ��Michael J. Behe���ڡ�Darwin��sBlack Box: The Biochemical Challenge to Evolution�������ģ�����ĵĺ�ϻ�ӡ��������۶Խ����۵���ս����˵��������ѧ��Ŭ����ʵ��ͼ�������֤�������״Ӻζ�����ʱ��������ѧ����ȴû�������κ�Ŭ��ͨ��ʵ��ͼ�����֤���Ӳ���ϵĽ����龰���������������ѧ��ͣ����50�������֧��������Դ�о�����ͬ��˼ά����С���Behe 1996�����ҵ����ñ�ϣ������������������ ������ѧ���ڡ��������������ղ������1999��˵�����ܹ��������һ�����ۣ��������ʵ��֤����������������֮�𡭡�û��ʵ��֤��֧���κ�һ�����ۣ�����Ϊ�����Ƶ��ܲ���˴��š������������ۻ����ҵĸ��ָ�����������Դ���۵���������Ҳ����û��һ������� �Һ���ͬ������ѧ�ң�Ҳ����ѧ�Һ�����ѧ�ң����ս��Ŀ����������л������������������ǵġ�����Ȼ����ʽʮ�ֵĶ࣬�ȿ��ﲻ֪�������������Ϊֻ���ɵ������ʵĺ�С���֣����µ���ѧֻ����Ϊ��֧�����л�������ǿ����ڵ������ʱ���ѿ�ʼ��һ��Ҳ��������Ϊ�١��������ڿ�ʼʱ����Ĵ��ڣ��������Ժ������ᷢ������ΪʲôҪ����ã��̫����Ѱ�ҵ������Դ�ء������Dz������������ڵĵ������������еĻ�ѧ��ɣ��Ȿ�������˾����������������������Դ����������ʱ����������أ����������ô˵�Ļ��������ˡ�ǰ�������ˣ������Ǵ��С�һ�������ġ���de Chardin 1955������Ȼ��������л���Ӧ���������ᡢ����������ʣ��������ࡢ�л���������Ļ����������ʡ� ���� �ڹ�ȥ�Ľ�һ�������У�����������Դ�����δͣϢ������Ҳ��δ��Ъ��������Դ����������һ�����������ļ��У�����ѧ˵Ʈ����ȥ��ǡ������������ɢ�Ļ�Ҷ���ɶ���������صġ����ݡ���Ȼ������ڻ������֮�С�������������ǽ������������Dz��ʲ�⧣��������˿�������һ��������Ϊ�Լ��Ŀ������������Ϊ��֤ʵ��Ҳ����֤α���������ã���ɣ���֪·�ж�Զ��ɽ�ж�ң����������ڡ���Ϊ�θ����������������һʫ��������1776�꣩�е�һ�����ԣ���һ���ն�����ʹ������æµҲ���Ҹ���һ������ʹ�����ĸе���ʵҲ���Ҹ�������ʷ��̨�ϵ�������Դ�ۣ�Ī������ˣ� �����Ƿ�Ӧ�����µ¹�ΰ����ѧ�Һڸ����Georg WilhelmFriedrich Hegel��1770��-1831�꣩�����ԣ����л��Ķ�������ʵ�Ķ��������ֶ����Լ�ά���Լ��������������ڵĹ��̣��������Լ����ձ鶫���������ձ鶫�����Լ�����Ϊ�Լ��ĸ������֡���Щ�����ڴ�������ʱ�����������Լ����� �����Ƿ�Ӧ����˼������ѧ�ҵ��ս���Pierre Teilhardde Chardin��1881�꣭1955�꣩�����һ����Ϊ����ṹ֮���������ĸ���������ڶ����Կ�����һ��������ʱ����ά�е�˲���ˡ�������֮�����ʸ�Ԫ�ؼ�����������ب�����������Ǹ�ֲ�������ȵĹ�ȥ����չ��ңԶ�첻�ɼ���δ��֮ϵ���е�һ���ְ��ˡ�����̽����ãã��ҹ֮�У�����ҵ�һյ��ҫ�����ƣ� ��Ҫ�ο��� лƽ. 2014. ��������Դ����������֮���������.��������ѧ�����磨Ӣ�ģ�Xie P. 2014. The Aufhebung and Breakthrough of the Theories on the Origin and Evolution of Life. Beijing: Science Press�� ���ߣ�лƽ ��Դ����ѧ�� �������㯣�����Դ�Ժη��� | 









 ������
������ �ö���
�ö��� ��Ĭ��
��Ĭ�� ������
������ ��ɫ��
��ɫ�� ǧ�ﶥ
ǧ�ﶥ ������
������
 ¥��
¥��
